|
|
Original Articles Indian Pediatrics 2005; 42:123-129 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
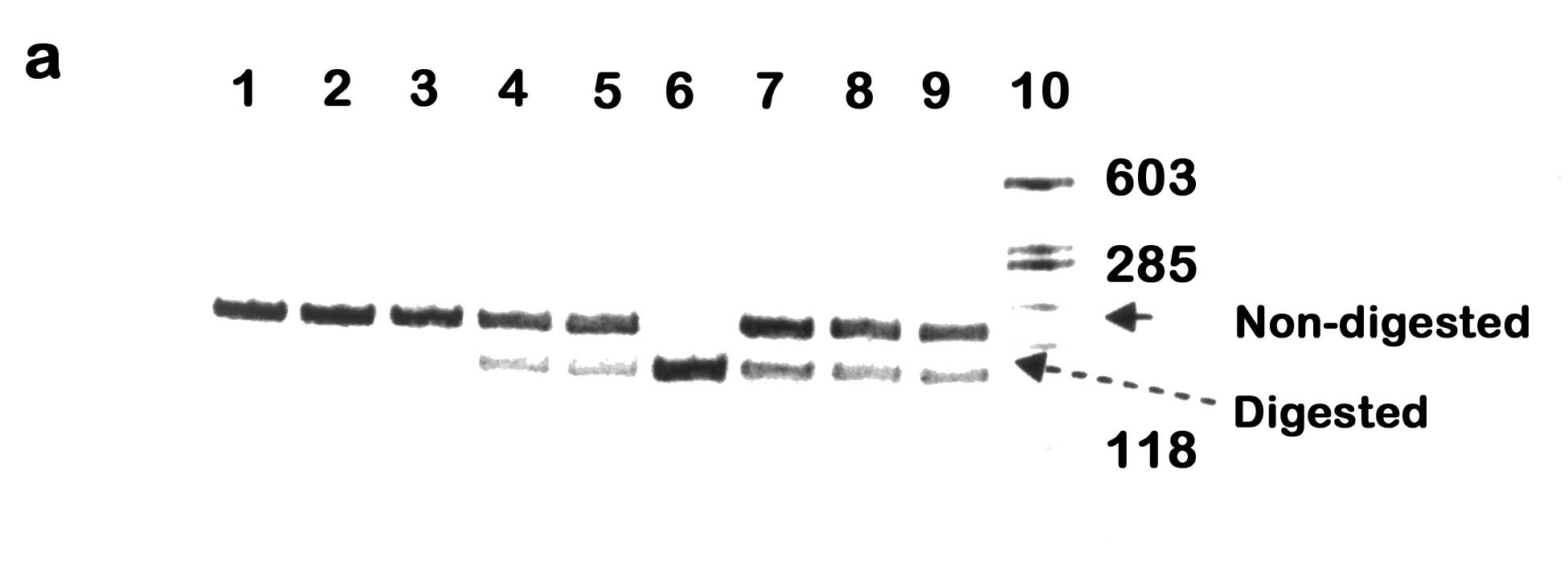
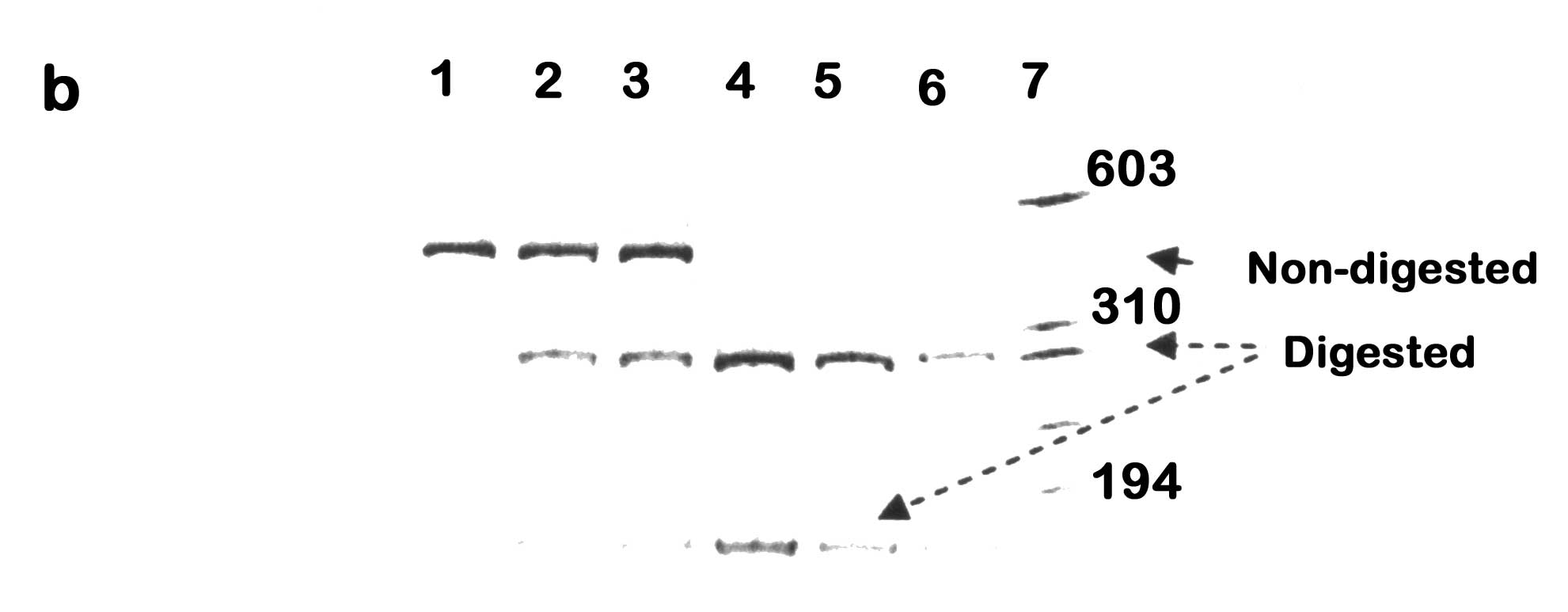
Abstract: To study the association of Attention Deficit Hyperactivity Disorder (ADHD) and polymorphism in the dopamine beta hydroxylase (DBH) gene in Indian ADHD cases. Subjects: Forty one ADHD cases were diagnosed as per the DSM-IV-TR criteria and evaluated by Conners’ Parents and Teachers Rating Scale and Wechsler’s Intelligence Scale for Children. Methods: Genomic DNA was amplified for exon 2 *444g/a and intron 5 (Taq I) polymorphism in the DBH gene followed by restriction fragment length polymorphism (RFLP) analysis. Haplotype-based haplotype relative risk (HHRR) was analyzed to ascertain the transmission pattern of these two polymorphisms in ADHD cases. Linkage disequilibrium (LD) between the two polymorphisms was calculated using EH+ and 2LD programs. Results: In the limited number of samples analyzed, a slight increase in transmission of the 444a allele in ADHD subjects was observed for DBH* 444g/a. The intron 5 (Taq I) polymorphism showed no significant association with ADHD in these cases. Strong disequilibrium was observed between DBH*444g/a and intron 5 (Taq I) polymorphism. Conclusion: This is the first molecular genetic study on ADHD in Indian subjects exploring transmission of polymorphisms in the DBH gene. Preliminary investigation shows a trend towards association between the transmission of DBH*444a allele and ADHD. No association was noticed between transmission of intron 5 (Taq I) polymorphism and ADHD in the Indian subjects. Presence of strong LD may point towards co-segregation of these two polymorphisms more often than expected. Attention Deficit Hyperactivity Disorder (ADHD) is a childhood onset neurobehavioral disorder showing strong genetic component(1). The prefrontal cortex of ADHD patients is found to be of smaller size in comparison to normal healthy individuals(2) and imaging studies revealed changes in the "fronto-subcortical" regions of the brain that are rich in dopaminergic innervations(3). Medications often prescribed to ADHD patients for symptomatic relief include psycho-stimulants like methyl-phenidate and amphetamine(4), which act primarily on the dopaminergic system. These observations led to the emphasis on dopamine (DA) as a target system to study the etiology of ADHD. Extensive molecular genetic studies on different ethnic groups have indicated that the gene for the enzyme dopamine beta hydroxylase (DbH) has a possible role in the etiology of ADHD(5-7). However, till date, a detailed genetic study of ADHD in Indian subjects is lacking. The metabolism of DA to norepinephrine (NE) is brought about by D βH, which arises from the sympathetic nervous system and is readily detectable in the plasma and cerebrospinal fluid(8). The structural gene encoding DβH (locus name DBH) is ~23kb long, contains 12 exons and is localized on chromosome 9q34(9). The gene is in tight linkage with the ABO locus and their oligogenic inheritance pattern is reported to be the cause for inter-individual variation observed in human serum DβH level(9,10). The repeated associative findings between abnormalities in the DA/NE systems and psychiatric abnormalities make DBH an important candidate gene to study neurobehavioral disorders like ADHD.Objective of the present preliminary study was to explore the possibility of association between Indian ADHD children/adolescents and two diallelic polymorphisms in the DBH gene. One is a single-base substitution at the 3' end of exon 2 named DBH*444 g/a, which is reported to be associated with altered CSF and plasma DβH activity in the European and African-American subjects(11,12). The other polymorphism is a Taq I restriction site polymorphism in the intron 5 of DBH that is found to be associated with ADHD in the Irish and Brazilian population(5-7). We sought to search for similar association between Indian ADHD subjects and polymorphisms in the DBH gene. Subjects and Methods ADHD cases were recruited from the Out-Patient Clinic of Manovikas Kendra Rehabilitation and Research Institute for the Handicapped in Kolkata. The age range of the probands was between 2.4 and 14 years, with 35 males and 6 females. The subjects were diagnosed as per the Diagnostic and Statistical Manual of Mental Disorders -IV-Text revised version (DSM-IV-TR, American Psychiatric Association), 2000(13). All available clinical information, prenatal history, developmental milestones and family records were collected by a team of neurologist, child psychiatrist, clinical psychologist and basic researchers. The subjects were also evaluated on the following scales: (1) The Conners’ Parents and Teachers Rating Scale and (2) Wechsler’s Intelligence Scale for Children (WISC) for the inattention-hyperactivity level and IQ status, respectively. Those suffering from any physical disability, psychiatric illness, pervasive developmental disorders, any form of mental retardation including fragile-X syndrome or with evidence of any other major medical or neurological conditions were excluded from the study. Peripheral blood was collected from the subjects and their parents, which includes 35 complete parent-offspring trios and 6 duos with only one parent, after obtaining informed written consent. DNA was extracted from leukocytes using the standard phenol/chloroform protocol(14). Genotyping PCR amplification was carried out for two polymorphisms of DBH gene, exon 2 *444 g/a polymorphism and intron 5 (Taq I) polymorphism(11). The primer sequence to amplify the 207-bp PCR fragment of exon 2 and surrounding intronic sequences were: sense, 5'-CCT GGA GCC CAG TGC TTG TC-3', and antisense, 5'-ACG CCC TCC TGG GTA CTC GC-3'(11). PCR amplification was performed at: initial denaturation at 95ºC for 7 min, 35 cycles of denaturation at 95ºC for 30 sec, annealing at 63ºC for 30 sec and extension at 72ºC for 45 sec. A final extension at 72ºC for 5 min was added. The 464-bp PCR fragment of intron 5 was amplified with the following primers: sense, 5'-CTG TAT TTG GAA CTT GGC ATC-3', and antisense, 5'-AGG CAT TTT ACT ACC CAG AGG-3'(7). PCR condition for intron 5 was: initial denaturation at 95ºC for 5 min, 35 cycles of denaturation at 94ºC for 1 min, annealing at 56ºC for 1 min and extension at 72ºC for 1.5 min, followed by a final extension at 72ºC for 5 min. PCR amplification for both polymorphisms were carried out using Perkin Elmer thermal cycler (Gene Amp #2400) in a final reaction volume of 40 µL containing 75-100ng of genomic DNA, 20 pmoles of each primer, 0.2 U Taq polymerase, 200 µM dNTP mix, 5% glycerol and 10 mM Tris buffer (Genei) with 50 mM KCl and 1.5 mM MgCl2. Restriction Fragment Length Polymorphism (RFLP) analysis DBH*444g/a in exon 2 is a silent polymorphism(12). DBH*444g (allele A1) is cleaved by the restriction endonuclease EcoN I yielding two fragments of 169 and 38-bp respectively(11); DBH*444a (allele A2), however, remains undigested. 5 µL PCR product was digested at 37ºC for 3 hrs. in a final volume of 25µL containing 1 U of EcoN I (New England Biolabs (NEB) and 1X strength NEB Buffer 4. The Taq I polymorphism in intron 5 of the DBH gene refers to a gain of restriction digestion site, which can be cleaved by the endonuclease Taq I(7). The A1 allele for this polymorphism is the undigested 464-bp PCR product, whereas, A2 allele, on digestion with Taq I, yields two fragments of 300 and 164-bp, respectively. 5 µL of the PCR product was digested at 65ºC for 2 hrs. in a final volume of 25 µL containing 2 U of Taq I (Genei), 1X Buffer B and 1X Bovine Serum Albumin. The digested products of both polymorphisms were separated by 12% polyacrylamide gel electrophoresis at 200 V for 3 hrs., stained with ethidium bromide and visualized using UV transilluminator. Statistics We used the Haplotype-based haplotype relative risk (HHRR) design to ascertain association between ADHD and transmission of genetic polymorphisms(15). This robust statistical method uses the non-transmitted parental alleles as ‘controls’ for evaluating transmission of allele and thus prevents spurious association due to population stratification. The EH+ program, version 1.2(16) was utilized to estimate the haplotype frequencies for the different alleles of DBH*444 g/a and intron 5 (Taq I) polymorphism. Subsequently, we used the 2LD program(17) to compute the disequilibria (D & D’) values for DBH*444 g/a and intron 5 (Taq I) polymorphism(18). Results Genotyping (Fig.1a) and HHRR analysis of DBH *444 g/a (Table I) in 41 Indian ADHD children and adolescents revealed that there is a minor increase in the transmission of allele 2 of the exon 2 EcoN I polymorphism (444 a) in ADHD subjects. However, the data is not statistically significant (X2 = 0.789, P = 0.3744, RR = 0.86). Genotyping (Fig.1b) and HHRR analysis of intron 5 (Taq I) polymorphism (Table I) suggest lack of significant association between transmission of this polymorphism and ADHD in the cases studied (X2 = 0.14, P = 0.7082, RR = 0.93).
TABLE I HHRR Analysis of DBH *444 g/a Poly-morphism and Intron 5 RFLP in 41 Nuclear Families with ADHD Probands.
* df = 1. The estimated haplotype frequencies for the alleles of the two polymorphisms were obtained from the EH+ program output (Table II). The 2LD program was used to compute this data for calculating LD between DBH*444 g/a and intron 5 (Taq I) polymorphism. Very strong LD was detected between these two polymorphisms (X2 = 43.31, df = 1, P < 0.0001). TABLE II Estimated Haplotype Frequencies (EH Fr) and Values of D and D’ for DBH *444 g/a - Int 5 RFLP Haplotypes in Indian ADHD Cases.*
*X2 for Disequilibria (D) = 43.31; df = 1; P < 0.0001. Discussion We have carried out a family-based association study on 41 nuclear Indian families having at least one ADHD child. The HHRR analysis has been used to predict the association between ADHD and transmission of exon 2*444g/a and intron 5 (Taq I) polymorphisms in the DBH gene. Significant association was reported earlier between DBH*444g/a and CSF DbH activity in European-American patients with mood or anxiety disorder(11). Recently, Hawi, et al(6) noticed a slight increase in the transmission of 444a allele in Irish ADHD cases (X2 = 0.9, P = 0.34, RR = 1.1), which was, however, not statistically significant. Our preliminary investigation on Indian ADHD cases also shows a small increase in transmission of the 444a allele, which is statistically in-significant. Lack of significant association was also noticed between transmission of the intron 5 (Taq I) polymorphism and ADHD in our study group. However, in Irish population, in contrast to our findings, significant association was observed between ADHD and Taq I A2 allele(6). Another report on Brazilian subjects showed similar association between the Taq I A2 allele and ADHD(5). On the contrary, a Milwaukee Longitudinal study in Caucasians found that the Taq I A1 allele is significantly associated with the disorder(19). Therefore, our data on the lack of association between any allele of intron 5 (Taq I) with ADHD may hint towards a unique genotypic pattern in our study group as compared to other ethnic groups. Further investigation employing large number of samples would enable us to rule out the possibility of association between these polymorphisms and ADHD in Indian subjects. Linkage disequilibrium analysis amongst DBH*444g/a and intron 5 (Taq I) polymorphism utilizing EH+ and 2LD programs was also carried out in the present study. DBH*444g/a has been reported to be in LD with several polymorphisms like DBH STR, intron 5 (Taq I) polymorphism and DBH*5'-ins/del(6,11,20). The present study also shows a very strong LD between DBH*444g/a and intron 5 (Taq I) polymorphism, suggesting that the disequilibrium pattern of the two polymorphisms are similar to other ethnic groups including Irish and European-American subjects(6,11). LD is a powerful alternative to linkage analysis for mapping and detecting genes with modest or minor effect and shows nonrandom association between alleles at different loci(21). As has been stated earlier, recombination history of the population within the particular segment of DNA is reflected from the presence of disequilibrium between two polymorphisms (22). However, LD can vary between different population due to recombination, genetic drift, population admixture and many such factors(23). Similar pattern of LD in Irish(6) and Indian population may point towards low frequency of recombination between these two polymorphisms which are co-segregated more often than expected. Lack of significant association between disease and transmission of different polymorphisms which are in disequilibria have been reported earlier in other ethnic groups (6). The present preliminary investigation on ADHD in Indian subjects revealed similar pattern of transmission. We are extending our study by using larger sample size and looking into more susceptibility loci for ADHD which would give us better understanding of the etiology of ADHD in Indian subjects. Acknowledgement Junior research fellowship to N. Bhaduri is gratefully acknowledged to the Council for Scientific and Industrial Research, Government of India. Thanks are also due to Dr. K. Chaudhuri, Scientist E II, Indian Institute of Chemical Biology, Kolkata and Dr. K. Nandagopal, Scientist, Manovikas Bio-medical Research and Diagnostic Center, Kolkata for their intellectual contribution. Contributors: NB was responsible for acquisition of data and analysis and drafting the article. SS performed psychometric analysis and clinical diagnosis, intellectual contribution. AC helped in psychological evaluation and IQ determination. PKG was responsible for neurological investigations and intellectual contribution. MS revised the manuscript critically. KM was responsible for concept and study design, interpretation of data and revising the article critically. Final manuscript was approved by all authors. Funding: None. Competing interests: None stated .
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
![]()